
Sarpa salpa (Linnaeus 1758) es el nombre de una especie de pez que se pesca y se comercializa con el nombre de salpa o salema. Se observó que durante algunas épocas del año, en las costas de Túnez, aparecían casos de efectos alucinógenos relacionadas con el consumo de este pescado. Sarpa se alimenta mayoritariamente de Posidonia oceanica A partir de ahí, se estudió el contenido estomacal de estos peces a lo largo del año para determinar la variación en su dieta. El estudio lo realizaron K. Bellassoued, A. Hamza, A. Abdelmouleh, J. van Pelt y A. Elfeki.
ESTE ES EL ARTÍCULO:
Respuesta antioxidante de una especie de pez herbívora (Sarpa salpa): Variación estacional
Antioxidant response of a common herbivorous fish species (Sarpa salpa): Seasonal variation
K Bellassoued1, 2*, A Hamza2, A Abdelmouleh2, J van Pelt3, A Elfeki1
1 Laboratory of Animal Ecophysiology, Science Faculty, University of Sfax, BP 1171, 3018 Sfax, Tunisia.
2 National institute of Sciences and Technologies of the Sea, Sfax, Tunisia.
3 Liver Research Facility/Laboratory of Hepatology, University Hospital Gasthuisberg, 3000 Leuven, Belgium. * Corresponding author. E–mail:khaledbra@yahoo.fr, khaledbra3@gmail.com
Received July 2010
Received in revisedform February 2011
Accepted May 2011
Received in revisedform February 2011
Accepted May 2011
RESUMEN
En el marco de un muestreo biológico de las especies marinas en las costas de Túnez, se estudió la salema Sarpa salpa, un pescado que es consumido por los habitantes de esta región. Se ha observado un acontecimiento estacional de efectos alucinógenos en los seres humanos después de consumir este pescado. El objetivo de este trabajo fue determinar la variación estacional de la composición de la dieta de S. salpa y correlacionar esta variación con la actividad antioxidante observada en los tejidos. La salema que se encuentra alrededor de la isla de Kerkennah es principalmente un pez herbívoro durante todas las estaciones del año. Se observó un incremento de la expresión de la catalasa (CAT) y la glutatión peroxidasa (GPx) en ciertos órganos en comparación con el pez control, Diplodus annularis. Se encontró un efecto acumulativo dependiente de la estación, que apareció primero en el hígado, después en el cerebro y por último en la carne. Además, se incrementa según el tamaño del animal y, por tanto, la cantidad de alimento que consume. El efecto alucinógeno que el consumo de la salema tiene en el ser humano se relaciona con la variación estacional de las epifitas que son coingeridas por estos peces con su comida. También se observó una correlación significativa (P < 0.01) entre el total de dinoflagelados tóxicos y la respuesta antioxidante: CAT y GPx en el hígado, el cerebro y la carne para todas las estaciones y tallas en conjunto.
Palabras clave: Sarpa salpa, dieta, dinoflagelados tóxicos, enzima antioxidante, catalasa, glutatión peroxidasa.
ABSTRACT
Within the framework of a biosurveillance of marine species on Tunisian coasts, we studied the salema Sarpa salpa, a fish that is consumed by the people living in this region. There is a seasonal occurrence of hallucinogen effects observed when this fish is consumed by humans. The aim of this work was to determine the seasonal variation in the composition of this fish's diet and to correlate this variation with the antioxidant activity found in its tissues. The salema that lives around the Island of Kerkennah is primarily a herbivorous fish during all seasons. We observed an increased expression of catalase (CAT) and glutathione peroxidase (GPx) in certain organs compared to the control fish, Diplodus annularis. There was a "season–depending" cumulative effect, appearing in the organs starting with the liver, followed by the brain, and finally the flesh. Moreover, it increases according to the size of the animal and thus to the amount of food it consumes. The hallucinogen effect that the consumption of salema has on humans is parallel to the seasonal variation in the epiphytes that are co–ingested by these fish with their food. A significant correlation (P < 0.01) was also observed between the total toxic dinoflagellates and the antioxidant response: CAT and GPx in liver, brain, and flesh for all seasons and all sizes together.
Key words: Sarpa salpa, toxic dinoflagellate, antioxidant enzyme, catalase, glutathione peroxidase.
INTRODUCCIÓN
La eutrofización de las aguas marinas costeras y el calentamiento global han causado una proliferación importante de microalgas y dinoflagelados particularmente tóxicos, los cuales producen una gran variedad de toxinas que contaminan los tejidos de los moluscos, los crustáceos y los peces. La concentración de la población humana en las zonas costeras ha aumentado la demanda de productos del mar y, por tanto, los casos de intoxicación por mariscos (Wu et al. 2005). La salema Sarpa salpa (Linnaeus 1758) es un pez herbívoro que se alimenta preferentemente de Posidonia oceanica durante todo el año (Peirano et al. 2001) y se utiliza para el consumo humano. Debido a su bajo costo, la salema aparece principalmente en el menú de la clase trabajadora de bajos ingresos. Se ha documentado la presencia de especies que causan la ciguatera y que viven como epifitas en las hojas de P. oceanica (Ben Brahim et al. 2010), las cuales son coingeridas por la salema como parte de su dieta (Velimirov 1984). Inclusive, el consumo de la salema no es recomendable en ciertos periodos del año (especialmente en verano y otoño) ya que causa un síndrome alucinatorio y trastornos del sistema nervioso central, que durante mucho tiempo fueron atribuidos a la presencia de caulerpina en el alga verde Caulerpa prolifera (Haro et al. 1994).
Las epifitas son importantes contribuyentes a la producción primaria de los ecosistemas de pastos marinos y presentan importantes fluctuaciones estacionales de la biomasa y la productividad (Mazzella et al. 1992). El crecimiento de las epifitas se ve influenciado por los mismos factores abióticos que afectan la planta huésped (e.g., irradiación, temperatura y nutrientes inorgánicos disueltos; Williams y Ruckelshaus 1993). Los factores bióticos también juegan un papel importante en la conformación de la comunidad epifítica, sobre todo en el cambio de hojas de los pastos marinos (Romero 1989) y el pastoreo (Neckles et al. 1993, Williams y Ruckelshaus 1993), ya que las epifitas algales suelen ser un alimento más atractivo para los herbívoros que las mismas hojas de los pastos marinos (Kitting et al. 1984). En este contexto, el golfo de Gabes (sur de Túnez, 35–33° N), que ha sido sujeto a presiones ambientales debido a actividades industriales y urbanas (Hamza–Chaffai et al. 1997, Zairi y Rouis 1999), ha experimentado una proliferación importante de microalgas y dinoflagelados particularmente tóxicos (Turki et al. 2006). Se ha demostrado que la proliferación de microalgas no deseadas es un problema creciente en ambientes tanto costeros como estuarinos (Smayda 1997, Leong y Taguchi 2005). Mar adentro en el golfo de Gabes, los aportes de nutrientes son influenciados por la circulación frontal y de agua atlántica y mediterránea (Estrada et al. 1985, Font et al. 1995). Esto se asocia con la degradación de las praderas marinas de P. oceanica y, consecuentemente, con la declinación de los recursos pesqueros en el golfo de Gabes.
La intoxicación alimentaria por ciguatera se debe a la presencia en el pescado de una toxina producida por el alga bentónica Gambierdiscus toxicus y otras microalgas coralinas, la mayoría pertenecientes a tres géneros:Prorocentrum, Ostreopsis y Amphidinum. Una forma menos común de ictiotoxismo es la intoxicación alucinógena(ichthyoallyeinotoxism), caracterizada por el desarrollo de trastornos del sistema nervioso central, especialmente alucinaciones y pesadillas. Se ha documentado este tipo de intoxicación en varios lugares del mundo (Halstead 1988, Chateau–Degat 2003) y es muy común en las zonas tropicales y templadas de los océanos Índico y Pacífico, así como en el mar Mediterráneo. Las especies ícticas descritas como alucinógenas en la literatura pertenecen a las siguientes ocho familias: Acanthuridae, Kyophosidae, Mugilidae, Mullidae, Pomacentridae, Serranidae, Siganidae y Sparidae. Sarpa salpa se alimenta principalmente de las hojas de P. oceanica, las cuales muestran un alto grado de colonización de epifitas (Peirano et al. 2001). Las hojas de P. oceanica y las epifitas que mantienen representan una fuente de alimento importante para las distintas especies que pastorean como el equino–dermo Paracentrotus lividus (Kirkman y Young 1981) y S. salpa, un pez espárido que forma cardúmenes (Velimirov 1984). Cabe señalar que la toxicidad de todas las especies varía según la zona de pesca, la temporada y el método de preparar el pescado para su consumo (Helfrich y Banner 1960). Las toxinas se incrementan a niveles peligrosos para los seres humanos durante su trasmisión mediante peces herbívoros y carnívoros, y varias especies resultan contaminadas (Vaillant et al. 2001).
El metabolismo oxidativo celular es una fuente continua de especies reactivas de oxígeno (ERO); producidas por la reducción univalente de O2, estas especies altamente reactivas pueden dañar la mayoría de los componentes celulares. El sistema antioxidante celular incluye enzimas como la superóxido dismutasa, la catalasa, la glutatión peroxidasa y la glutatión reductasa que actúan mediante la detoxificación de las ERO generadas. No obstante, en muchos casos, la tasa de generación de ERO excede la de su remoción, provocando el estrés oxidativo e incrementando los marcadores oxidativos. Es bien conocido que los contaminantes, tales como varios pesticidas (Sayeed et al. 2003) y metales (Almeida et al. 2002), así como la exposición hiperóxida (Lushchak y Bagnyukoca 2006) estimulan mecanismos de protección contra ERO. En un ambiente natural como el mar, existe la intoxicación de organismos acuáticos por especies de fitoplancton tóxicas. Por ende, es importante estudiar el estrés oxidativo en peces debido a los posibles efectos sobre los organismos y en relación con el consumo de pescado por los seres humanos.
Los contaminantes en organismos acuáticos incrementan la producción de ERO, provocando daño oxidativo (Shiet al. 2005, Livingstone 2001). Se han propuesto los cambios en los niveles de defensa antioxidante endógena como marcadores biológicos de la contaminación acuática (Camus et al. 2004, Livingstone 2001). La activación del sistema enzimático antioxidante sugiere que el sistema de defensa juega un papel importante en la respuesta a una elevada situación oxidativa. La determinación de las actividades de las enzimas antioxidantes en los órganos de peces permite detectar la presencia de contaminantes y toxinas (Sureda et al. 2006), tales como la ciguatoxina, que se presenta en las epifitas de las praderas de P. oceanica. El objetivo de este trabajo fue especificar los patrones estacionales de las fuentes de alimento y las actividades antioxidantes en el hígado, el cerebro y la carne de S. salpa y correlacionarlos.
MATERIALES Y MÉTODOS
Recolección y muestreo
Este estudio se realizó en las inmediaciones de la isla de Kerkennah (golfo de Gabes, sureste de Túnez). Este archipiélago se caracteriza por extensas praderas de P. oceanica. Los especímenes de S. salpa se recolectaron entre enero de 2006 y enero de 2007, en diferentes momentos de cada estación del año. Se recolectaron 59 especímenes en invierno, 57 en primavera, 57 en verano y 55 en otoño. Las tallas variaron entre 12.8 y 28 cm. A cada organismo se le midió la longitud total (LT) con una precisión de 0.1 cm y el peso total con una precisión de 0.1 g. Inmediatamente después de su captura, los peces fueron disecados y los intestinos fueron retirados y preservados en una solución de formalina al 4%. En el laboratorio, las presas se identificaron hasta el nivel taxonómico más bajo posible. La abundancia de las especies y el peso húmedo se registraron con una precisión de ±0.001 g, después de eliminar el agua superficial con un papel secante. Asimismo, inmediatamente se retiraron el hígado, el cerebro y la carne, y se lavaron con una solución salina helada y se conservaron a –30 °C en las mismas condiciones en las que se mantuvo el pez control, el raspallón Diplodus annularis (Linnaeus 1758), que pertenece al mismo biotopo y a la misma familia.
Análisis del contenido estomacal
Para evaluar los posibles cambios en la dieta en relación con la talla, los individuos se dividieron en dos clases de talla según Pallaoro et al. (2008): adultos (3–7 años), LT > 20 cm (talla grande); y subadultos (2 años), 17 cm < LT < 20 cm (talla mediana).
Se analizó la composición y fuente de la dieta de cada espécimen cuyo estómago estaba lleno de hojas y epifitas. El contenido estomacal se lavó en una caja de Petri y se examinó debajo de un microscopio. Los componentes tróficos se clasificaron en grandes grupos taxonómicos y, cuando fue posible, se identificaron hasta nivel de especie de acuerdo con Fischer et al. (1987). La dieta de la salema se caracterizó usando el índice de vacuidad (IV), la masa relativa del componente i y la composición gravimétrica (% Cp, el peso húmedo de los componentes en el peso húmedo total de los estomágos no vacíos), según las siguientes fórmulas:

El análisis del índice de vacuidad a través del tiempo nos proporciona información del comportamiento alimentario y nos permite obtener ritmos estacionales o mensuales.
El componente i puede ser un grupo, una familia, un género o una especie según el grado de determinación.
Análisis de las especies microepifíticas en el contenido estomacal
El contenido estomacal se introdujo en un frasco al cual se añadieron 500 mL de agua de mar. El frasco se agitó y la mezcla se filtró. El agua filtrada se preservó en formol al 3%.
El fitoplancton se identificó a partir de células vivas para evitar la destrucción de células. La enumeración de las células se realizó con un microscopio invertido, siguiendo el método propuesto por Utermõhl (1958), después de la fijación con una solución de yodo (Lugol) al 4% (Bourrelly 1985). Las muestras fitoplanctónicas se identificaron de acuerdo con Tregouboff y Rose (1957), Huber–Pestalozzi (1968), Dodge (1973, 1975, 1982, 1985), Balech (1988, 1995) y Tomas et al. (1993, 1996). Se siguió el mismo protocolo en las cuatro estaciones (invierno, primavera, verano y otoño).
Se registró la concentración de células fitoplanctónicas (diatomeas no tóxicas) y de dinoflagelados epifitos tóxicos asociados con la intoxicación por ciguatera (e.g., Prorocentrum sp., Ostreopsis sp., Coolia sp. yAmphidinum sp.).
Preparación de los extractos citosólicos
Las muestras congeladas de las células del hígado, el cerebro y la carne se homogeneizaron (Ultra Turrax T25, Alemania; 1:2, p/v) en un tampón helado (TBS: 50 mM Tris, 150 mM NaCl, pH 7.4) y se centrifugaron (5000 g, 30 min, 4 °C). Los sobrenadantes se congelaron a –30 °C.
Ensayos bioquímicos
En los tejidos, se midió la actividad enzimática de la catalasa mediante el método colorimétrico (con luz UV) descrito por Aebi (1984), usando H2O2 como sustrato; la actividad de la glutatión peroxidasa (GSH–Px) se midió mediante una modificación del método colorimétrico propuesto por Flohe y Günzler (1984), usando H2O2 como sustrato junto con una reducción de GSH. El contenido proteico de los extractos de tejido se determinó de acuerdo con Bradford (1976).
Análisis estadístico
Los datos se presentan como medias ± desviación estándar. Las determinaciones se realizaron a partir de cinco animales por grupo y las diferencias se analizaron mediante un análisis de varianza de dos vías (factores fijos: talla y estación), seguido por la prueba de Fisher (Stat View); el nivel de significancia se fijó en P < 0.05. Asimismo, se calcularon coeficientes de correlación (R) para todas las tallas y estaciones en conjunto usando la correlacion de Pearson.
RESULTADOS
Contenido estomacal de la salema
Durante el año de estudio, la dieta de adultos y subadultos de S. salpa consistió de una variedad de flora marina (tabla 1). A pesar de la abundancia de Caulerpa prolifera en el biotopo, esta especie no apareció en los contenidos estomacales.

Índice de vacuidad
Para el análisis del comportamiento alimentario de S. salpa se examinaron 228 estómagos, de los cuales 104 contenían especies de macrófitos. Los peces se capturaron alrededor de la isla de Kerkennah (golfo de Gabes) durante la noche, ya que es cuando generalmente se están alimentando y están más activos. Los individuos capturados se trasladaron a bordo la mañana siguiente, por lo que algunos podrían haber permanecido en la red durante varias horas y su captura pudo haber sucedido antes de la ingesta de presas o después de la digestión. Consecuentemente, varios de los especímenes tenían el estómago vacío al momento de su recolección.
La variabilidad estacional del índice de vacuidad promedio para las clases de talla grande y mediana mostró un decremento en primavera (31.57%) y verano (40%) en relación con otoño (66.10%), lo que indica que S. salpa se alimentó más en los primeros dos periodos.
Coeficiente de variación estacional de la masa (%) para Posidonia oceanica
La composición gravimétrica (%) del pastoreo de las hojas de P. oceanica por S. salpa incrementó gradualmente de invierno a otoño: 18.94% en invierno, 48.95% en primavera, 59.92% en verano y 74.8% en otoño. Esto muestra que las hojas de P. oceanica son el alimento preferencial de la salema en casi todas las estaciones. Las algas son un alimento complementario según su disponibilidad en el ambiente. El epibionte es un alimento ocasional o complementario para una clase de talla en casos extremos. La dieta no sólo está condicionada por la disponibilidad de alimentos, sino también por el comportamiento territorial y de búsqueda de refugio de la especie.
Dieta de la salema
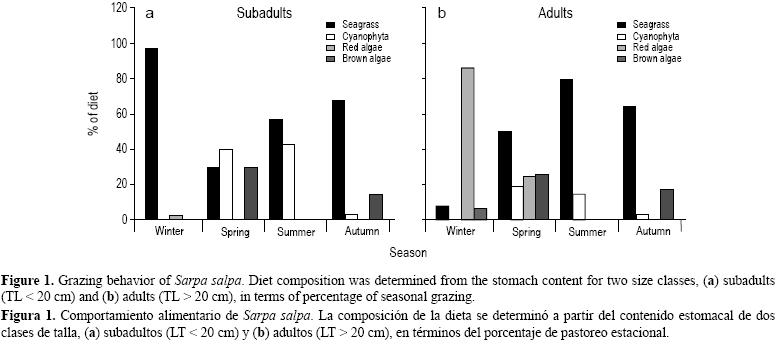
En invierno, los adultos de S. salpa consumieron menor cantidad de hojas de P. oceanica, constituyendo 7.34% de la dieta, e incrementaron su consumo de algas rojas (87.18%) y pardas (5.77%) (fig. 1). A diferencia de los adultos, para los subadultos se registró un pastoreo estacional de P. oceanica de casi 97.34% (fig. 1).
En primavera, el coeficiente del índice de vacuidad fue menor ya que el animal tiende a comer más. Las tasas de pastoreo de las hojas de P. oceanica fueron bastante altas para las dos clases de talla. Los peces de talla mediana incluyeron a Lyngbia sp. en su dieta (hasta 40%), ciertamente no por motivos preferenciales sino más bien debido a la abundancia de esta alga en el medio ambiente (fig. 1).
{kind=link}
El verano es la estación que precede el periodo reproductivo de los peces de talla grande. Los adultos incrementaron su consumo de las hojas de P. oceanica, alcanzando 79.5% durante esta temporada, mientras que los subadultos incluyeron un mayor porcentaje de Lyngbia sp. en su dieta, y el porcentaje de Lyngbia sp. (42.18%) fue significativo (fig. 1).
En otoño, el periodo reproductivo de esta especie, el animal no consume lo suficiente; sin embargo, se observó que su espectro trófico resultó muy diversificado por una gran cantidad de algas. El alga parda Cytoseira sp. (4.24%) sólo se identificó en esta estación. Tanto los adultos como los subadultos alcanzaron su máximo pastoreo de las hojas de P. oceanica (64.31% y 67.20%, respectivamente) (fig. 1).
Evaluación de la composición fitoplanctónica
El fitoplancton evaluado correspondió a la microflora epifítica algal digerida por S. salpa. La variación estacional del fitoplancton tóxico se evaluó en función del número total de fitoplancton. Las especies de fitoplancton tóxico en los contenidos estomacales fueron los dinoflagelados Prorocentrum mexicanum, Prorocentrum lima, Prorocentrum concavum, Ostreopsis siamensis, Coolia monotis y Amphidinum carterae. Las proporciones de las especies tóxicas incrementaron en conjunto con la población total de fitoplancton (fig. 2).

Variación estacional de las especies que causan ciguatera
Para evaluar el posible efecto de la toxicidad de las especies asociadas con la ciguatera en peces y, consecuentemente, sus consumidores, se observó que la variación estacional media del fitoplancton tóxico incrementó significativamente (P < 0.05) en el contenido estomacal de la salema durante el verano en comparación con el otoño, el invierno y la primavera (fig. 2). La cantidad de especies ciguatéricas coingeridas porS. salpa como parte de su dieta incrementó gradualmente de la primavera al verano, y se supone que se acumulan en los órganos de esta especie (fig. 2). Por tal razón, el consumo de la salema no es recomendable en ciertos periodos del año (especialmente en verano y otoño) ya que causa un síndrome alucinatorio y trastornos del sistema nervioso central.
Variación estacional de la actividad antioxidante en la salema
Se estudió la actividad de las enzimas antioxidantes en varios órganos y compartimientos de los especímenes deS. salpa examinados. Los resultados se compararon con los obtenidos para el pez control, Diplodus annularis (del mismo biotopo). Esto nos permitió observar varias especificidades.
En invierno, no se observó una variación significativa (P > 0.05) para la actividad antioxidante de la catalasa (CAT) y la glutatión peroxidasa (GPx) en los adultos y subadultos de S. salpa así como en D. annularis (figs. 3, 4).
En primavera, se observó un incremento significativo (P < 0.05) de la actividad antioxidante de CAT y GPx, el cual afectó sólo el hígado de los adultos y subadultos de S. salpa (figs. 3, 4).
En verano, se observó un aumento significativo (P < 0.05) de la actividad de CAT en el hígado de los adultos y subadultos de S. salpa. También se registró una variación significativa (P < 0.05) de la actividad de GPx en el hígado de ambas clases de talla, así como una variación significativa (P < 0.05) de las actividades de CAT y GPx en el cerebro de sólo los peces de talla grande (figs. 3, 4).
En otoñó, se observó una variación significativa (P < 0.05) de la actividad de CAT en el hígado de los peces de talla grande y mediana. También se registró una variación significativa (P < 0.05) en la actividad enzimática antioxidante en el cerebro y la carne de los adultos de S. salpa. La actividad de GPx se observó en los tres órganos de los adultos de S. salpa (P < 0.05) (figs. 3, 4).
Correlación interestacional entre el porcentaje de especies ciguatéricas y los perfiles de estrés oxidativo para la salema
A fin de justificar y consolidar nuestras observaciones, se realizó una serie de pruebas para determinar la correlación entre el total de dinoflagelados tóxicos y los perfiles del estrés oxidativo en los diferentes órganos examinados para todas las estaciones y todas las tallas en conjunto. Se observó una correlación significativa (P < 0.01) entre el total de dinoflagelados y las actividades antioxidantes de CAT y GPx en el hígado, el cerebro y la carne para todas las estaciones y tallas en conjunto (tabla 2).
DISCUSIÓN Y CONCLUSIONES
El índice de vacuidad promedio para S. salpa (54.38%) fue algo débil a lo largo del año, mientras que para D. annularis se registró un coeficiente de vacuidad promedio bastante alto: 91.48% (Derbal et al. 2007). Además, se observó que tal coeficiente decreció en primavera y verano; consecuentemente, S. salpa consumió más alimento en estas dos estaciones. El índice de vacuidad fue bajo en el periodo antes de la puesta de huevos, lo que indica que la salema se alimentó y almacenó lípidos para el proceso de maduración de los productos genitales. La salema presentó un periodo de desove máximo, de mediados de septiembre a mediados de octubre. El periodo de asentamiento intensivo se observó a finales de noviembre. Estos resultados son comparables con los obtenidos por Quignard et al. (1984).
Durante el verano, a diferencia del invierno, S. salpa aumentó su pastoreo de las hojas de P. oceanica,alcanzando un porcentaje de 79.65%, una tasa comparable con los resultados obtenidos por Alcoverro et al.(1995). En otoño, la mayoría de los especímenes de las dos clases de talla consumieron hojas de P. oceanica, por lo que este alimento fue el mayor componente en los contenidos estomacales de la salema (>50% en otoño). Por tanto, a pesar de la regresión en las últimas décadas, la pradera está en equilibrio con el medio ambiente y P. oceanica muestra un ciclo regular de crecimiento así como de la sucesión de colonización epifítica. Las hojas adultas, que mostraron mayor colonización por epifitas, fueron preferentemente consumidas por los herbívoros durante todo el año en todas las profundidades. De junio a septiembre S. salpa realiza el pastoreo en grandes cardúmenes que se alimentan activamente de P. oceanica a fin de acumular reservas para el periodo invernal cuando comen menos y de preparar los peces adultos para la reproducción. En marzo, cuando los adultos de S. salpa emigran a aguas profundas, los jóvenes habitan y se alimentan en fondos rocosos y poco profundos (Peirano et al. 2001).
Se observó que, junto con las algas y Posidonia, los dinoflagelados tóxicos presentan una gran diversidad y notables fluctuaciones estacionales. Además de las diatomeas, las especies de dinoflagelados tóxicos fueron identificadas como Prorocentrum mexicanum, P. concavum, P. lima, Ostreopsis siamensis, Coolia monotis yAmphidinum carterae. Prorocentrum mexicanum produce toxinas hemolíticas (Nakajima et al. 1981) y toxinas solubles en agua de acción rápida (Tindall et al. 1989). Prorocentrum concavum produce tres ésteres diólicos del ácido okadaico (Hu et al. 1993) y una toxina de acción rápida, y se le acusa de causar el síndrome de ciguatera.Prorocentrum lima produce toxinas diarreicas (intoxicación diarreica por mariscos) (Bomber y Aikman 1989), el síndrome de ciguatera (Faust 1991) y toxinas de acción rápida. Ostreopsis siamensis y Coolia monotis son especies que causan la ciguatera (Steidinger y Baden 1984). Esto indica que existen los dinoflagelados tóxicos y sustenta los resultados obtenidos por Raikhlin–Eisenkraft et al. (1988). Las proporciones de las especies tóxicas incrementaron en la misma dirección que la población total de fitoplancton. El dinoflagelado al nivel de las hojas de Posidonia oceanica adquiere mucha importancia a finales de primavera y en verano. Estas especies también fueron los dinoflagelados más abundantes durante el verano de 2001 en la columna de agua del lago de Bizerte (costa norte de Túnez) (Turki et al. 2006). Este aumento se debe principalmente al aumento de la superficie foliar, como resultado del crecimiento de las hojas en este periodo (Alcoverro et al. 1995). De hecho, el consumo de S. salpa no es recomendable en ciertos periodos del año (especialmente en verano y otoño) ya que causa un síndrome alucinatorio y trastornos del sistema nervioso central, los cuales por mucho tiempo fueron atribuidos a la presencia de caulerpina en el alga verde Caulerpa prolifera (Haro et al. 1994). Esta suposición no fue confirmada hasta hace poco ya que rara vez se detecta tal alga en el contenido estomacal de peces. Por tanto, según el presente estudio, las especies ciguatéricas son las causantes más que la caulerpina.
Los radicales oxidativos se generan continuamente como resultado del metabolismo normal y pueden aumentar como productos secundarios de las reacciones de biotransformación de las toxinas o los xenobióticos. Los sistemas antioxidantes celulares han demostrado una gran adaptación al estrés oxidativo para contrarrestar la producción excesiva de ERO (Alvarez y Boveris 1993, Niwa et al. 1996, Sureda et al. 2004). Se utiliza el estatus antioxidante de la célula a fin de evaluar la capacidad de los organismos para resistir el estrés ambiental inducido por algunos contaminantes marinos (Frenzilli et al. 2004).
En invierno, no se observó una variación significativa de la actividad de las enzimas antioxidantes en los diferentes órganos examinados de S. salpa en comparación con el pez control, D. annularis. Pareciera ser que esta ausencia de una variación significativa se debe a la ausencia de dinoflagelados tóxicos en las hojas de P. oceanica consumidas por S. salpa. En primavera, se observó un aumento significativo en las actividades de CAT y GPx en el hígado de la salema. El aumento en el porcentaje de P. oceanica como fuente alimentaria de los adultos y subadultos podría explicar la diferencia encontrada entre las respuestas de las enzimas antioxidantes a nivel hepático en las dos clases de talla. Además, se observó un aumento en la actividad de las enzimas antioxidantes que ciertamente se debe a la presencia de dinoflagelados tóxicos en el contenido estomacal de S. salpa con un porcentaje igual a 1.14%. Las respuestas de la actividad de CAT y de la superóxido dismutasa frecuentemente tienen perfiles similares; sin embargo, el aumento de la actividad de CAT es más fuerte y sucede antes que la de la superóxido dismutasa (Di Giulio et al. 1993).
En verano, las hojas de P. oceanica mostraron una mayor colonización de epifitas debido a que el agua circundante estaba más tranquila. Los periodos de máximo consumo de las hojas de P. oceanica por S. salpa se registraron en verano y otoño. Esto afecta las actividades antioxidantes resultantes. Asimismo, se observó una variación significativa de las actividades de CAT y GPx en el hígado de los peces de talla grande y mediana. Esto demuestra que la CAT principal es altamente expresada durante el periodo de contaminación por fitoplancton tóxico debido a la eliminación de ERO, lo cual concuerda con los resultados obtenidos por Di Giulio et al. (1993). También se observó una variación significativa de las actividades de CAT y GPx en el cerebro de los adultos de S. salpa.
En otoño, el porcentaje de especies asociadas con la ciguatera fue más importante (5.26%) que en las otras temporadas. Durante el otoño, los peces de ambas clases de talla consumieron las hojas de P. oceanica como la fuente de alimento preferencial (>50%), lo cual concuerda con los resultados de Peirano et al. (2001). Las diferencias en pastoreo pueden ser explicadas por un comportamiento diferente de los herbívoros. Los cardúmenes masivos de S. salpa se alimentan activamente de las hojas de P. oceanica durante el verano a fin de acumular reservas para la reproducción durante el otoño. Este comportamiento se refleja en la tasa de actividad antioxidante, y ambas clases de talla mostraron una variación significativa de la actividad hepática de CAT.
Las actividades de GPx se expresaron en los tres órganos de los adultos de S. salpa. Estos resultados nos permitieron concluir que hubo un pico en la toxicidad en otoño, y que la toxicidad aumentó notablemente en el cerebro en otoño e inclusive se acumuló en la carne de los peces de talla grande. El metabolismo de compuestos tóxicos con frecuencia resulta en la formación de ERO, lo cual contribuye a su toxicidad (Chovanec et al. 2003).
La inducción del aumento de la respuesta antioxidante es una respuesta lógica a la exposición a sustancias tóxicas generadas por los dinoflagelados. En este contexto, el fitoplancton es responsable de la activación de la actividad antioxidante o actúa en paralelo con otros factores de estrés como la influencia estacional y la saturación de oxígeno y la temperatura del agua. Estos factores pueden intervenir, pero el fitoplancton es el más influyente debido a los dinoflagelados tóxicos que consume la salema. El pez control, D. annularis, no consume dinoflagelados tóxicos y no presenta una respuesta antioxidante como S. salpa. Pareciera ser que las actividades antioxidantes se incrementan en presencia de fitoplancton tóxico en el contenido estomacal de la salema.
Se observó una correlación significativa (P < 0.01) entre el total de dinoflagelados tóxicos y las actividades antioxidantes de CAT y GPx en el hígado, el cerebro y la carne para todas las estaciones y tallas en conjunto. Por tanto, los efectos de las especies ciguatéricas inducen la actividad antioxidante, la cual comienza a incrementar durante la primavera y a decrecer durante el invierno. Estos resultados confirman lo documentado por Rodríguez–Ariza et al. (1993), quienes observaron un aumento paralelo de GPx, CAT y superóxido dismutasa en Mugil sp. contaminado.
En resumen, se ha mostrado que las actividades antioxidantes en S. salpa de la isla de Kerkennah presentan un efecto acumulativo dependiente de la temporada, apareciendo primero en el hígado, luego en el cerebro y por último en la carne. Este efecto aumenta con el tamaño del animal y, por ende, posiblemente con la cantidad de hojas de P. oceanica consumidas, sobre las que abundan epifitas fitoplanctónicas según la estación del año.
AGRADECIMIENTOS
Este trabajo fue apoyado por la unidad de investigación "Estrés Oxidativo y Salud" encabezado por A El Feki, profesor de la Facultad de Ciencias de la Universidad de Sfax (Túnez). Los autores agradecen a A Rebai (Centro de Biotecnología, Sfax, Túnez).
REFERENCIAS
Alcoverro T, Durate CM, Romero, J. 1995. Annual growth dynamics of Posidonia oceanica: Contribution of large–scale versus local factors to seasonality. Mar. Ecol. Prog. Ser. 120: 203–210. [ Links ]
Almeida JA, Diniz YS, Marques SF, Faine LA, Ribas BO, Burneiko RC, Novelli EL. 2002. The use of the oxidative stress responses as biomarkers in Nile tilapia (Oreochromis niloticus) exposed to in vivo cadmium contamination. Environ. Int. 27: 673–679. [ Links ]
Alvarez S, Boveris A. 1993. Induction of antioxidant enzymes and D–diaphorase in human blood mononuclear cells by light stress. Arch. Biochem. Biophys. 305: 247–251. [ Links ]
Balech E. 1988. Los dinoflagelados del Atlántico sudoccidental. Publicaciones Especiales del Instituto Español de Oceanografia, Madrid, 310 pp. [ Links ]
Balech E. 1995. The genus Alexandrium Halim (Dinoflagellata). Special publication, Sherkin Island Marine Station, Cork. [ Links ]
Ben Brahim M, Hamza I, Hannechi M, Rebai A, Jarboui O, Bouain A, Aleya L. 2010. Variability in the structure of epiphytic assemblages of Posidonia oceanic in relation to human interferences in the Gulf of Gabes, Tunisia. Mar. Environ. Res. 70: 411–421. [ Links ]
Bourrelly P. 1985. Les Algues d'Eau Douce. Initiation à la Systèmatique. Tome II. Les Algues Bleues et Rouges. Les Euglénins, Peridiniens et Cryptomonadines. Société Nouvelle des Editions Boubée, 57 pp. [ Links ]
Bradford M. 1976. A rapid and sensitive method for the quantisation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem. 7: 248–254. [ Links ]
Camus L, Pampanin DM, Volpato E, Delaney E, Sanni S, Nasci C. 2004. Total oxyradical scavenging capacity responses in Mytilus galloprovincialis transplanted into the Venice Lagoon (Italy) to measure the biological impact of anthropogenic activities. Mar. Pollut. Bull. 49: 801–808. [ Links ]
Château–Degat ML. 2003. Les toxines marines: Problèmes de santé en émergence. Vertigo 4: 1–11. [ Links ]
Chovanec A, Hofer R, Schiemer F. 2003. Fish as bioindicators. In: Markert BA, Breure AM, Zechmeister HG (eds.), Bioindicators and Biomonitors. Elsevier Science, Amsterdam. [ Links ]
Derbal F, Nouacer S, Kara MH. 2007. Composition et variations du regime alimentaire du sparaillon Diplodus annularis (Sparidae) du Golf d'Annaba. Cybium 31: 443–450. [ Links ]
Di Giulio RT, Habig C, Gallagher EP. 1993. Effects of rock harbour sediments on indices of biotransformation, oxidative stress, and DNA integrity in channel catfish. Aquat. Toxicol. 16: 311–320. [ Links ]
Dodge JD. 1975. The Prorocentrales (Dinophyceae). II . Revision of taxonomy within the genus Prorocentrum.Bot. J. Linn. Soc. 71: 103–125. [ Links ]
Dodge JD. 1982. Marine Dinoflagellates of the British Isles. Her Majesty's Statonery Office, London, 303 pp. [ Links ]
Dodge JD. 1985. Atlas of Dinoflagellates. A Scanning Electron Microscope Survey. Ferrand Press, London, 119 pp. [ Links ]
Estrada M, Vives F, Alcarez M. 1985. Life and productivity of the open sea. In: Marglef R (eds.), Western Mediterranean. Pergamon Press, Oxford, pp. 150–200. [ Links ]
Faust MA. 1991. Morphology of ciguatera–causing Prorocentrum lima (Pyrrophyta) from widely differing sites. J. Phycol. 27: 642–648. [ Links ]
Fischer W, Bauchot ML, Schneider M. 1987. Identification index of species for fishing needs. Review 1399, Mediterranean and black sea. Volume I. plants and aquatic Vertebrates. Pub. FAO Project GCP/INT422/EEC, Rome, 760. [ Links ]
Flohe L, Günzler WA. 1984. Assays of glutathione peroxidase. Methods Enzymol. 105: 114–121. [ Links ]
Font J, Garcia–Ladona E, Gorriz EG. 1995. The seasonality of mesoscale motion in the Northern Current of the western Mediterranean: several years of evidence. Oceanol. Acta. 18: 207–219. [ Links ]
Frenzilli G, Bocchetti B, Pagliarecci R, Nigro MM, Annarumma F, Scarcelli V, Fattorini D, Regoli F. 2004. Time–course evaluation of ROS mediated toxicity in mussels, Mytilus galloprovincialis, during a field translocation experiment. Mar. Environ. Res. 58: 609–613. [ Links ]
Halstead BW. 1988. Poisonous and Venomous Marine Animals of the World. Darwin Press, Princetown, New Jersey, 1168 pp. [ Links ]
Hamza–Chaffai A, Amiard–Triquet C, El Abed A. 1997. Metallothionein–like protein, is it an efficient biomarker of metal contamination? A case study based on fish from the Tunisian coast. Arch. Environ. Contam. Toxicol. 33: 53–62. [ Links ]
Haro L, Jouglard DE, Thomas MJ, David JM. 1994. Intoxications de type ciguatérique aprés ingestion de Sparidae de Méditerranée. In: Boudoresque CF, Meinsez A, Gravez V (eds.), First International Workshop on Caulerpa taxifolia. GIS Posidonie Publ. France, pp. 271–274. [ Links ]
Hu TA, De Freitas SW, Doyle J, Jackson D, Marr J, Nixon E, Pleasances S, Quilliam MA, Walter JA, Wright JLC. 1993. New DSP toxin derivatives isolated from toxic mussels and the dinoflagellates Prorocentrum lima andconcavum. In: Smayda T, Shimizu Y (eds.), Toxic Phytoplankton in the Sea. Elsevier, Amesterdam, pp. 507–512. [ Links ]
Huber–Pestalozzi G. 1968. Das Phytoplankton des Susswassars. 1. Halfte, Cryptophyceae, Chloromonadophyceae, Dinophyceae. E. Schweizerbart Verlag, Stuttgart, 322 pp. [ Links ]
Kirkman H, Young PC. 1981. Measurement of health and echinoderm grazing on Posidonia oceanica (L.) Delile. Aquat. Bot. 10: 329–338. [ Links ]
Kitting CL, Fry B, Morgan MD. 1984. Detection of inconspicuous epiphytic algae supporting food webs in seagrass meadows. Oecologia (Berlin), 62: 145–149. [ Links ]
Leong SCY, Taguchi S. 2005. Optical characteristics of the harmful dinoflagellate Alexandrium tamarense in response to different nitrogen sources. Harmful Algae 4: 211–219. [ Links ]
Livingstone DR. 2001. Contaminant–stimulated reactive oxygen species production and oxidative damage in aquatic organisms. Mar. Pollut. Bull. 42: 656–666. [ Links ]
Lushchak VI, Bagnyukoca TV. 2006. Effects of different environ–mental oxygen levels on free radical processes in fish. Comp. Biochem. Physiol. B: Biochem. Mol. Biol. 144: 283–289. [ Links ]
Mazzella L, Buia MC, Gambi MC, Lorenti M, Russo GF, Scipione MB, Zupo V. 1992. Plant–animal trophic relationships in the Posidonia oceanica ecosystem of the Mediterranean Sea: A review. In: John DM, Hawkins SJ, Price JH (eds.), Plant–Animal Interactions in the Marine Benthos. Clarendon Press, Oxford, pp. 165–187. [ Links ]
Nakajima I, Oshima Y, Yasumoto T. 1981. Toxicity of benthic dinoflagellates in Kinawa. Bull. Jap. Soc. Sci. Fish. 47: 1029–1033. [ Links ]
Neckles HA, Wetzel RL, Orth RJ. 1993. Relative effects of nutrient enrichment and grazing on epiphyte–macrophyte (Zostera marina L.) dynamics. Oecologia (Berlin) 93: 285–295. [ Links ]
Niwa Y, Ozaki T, Kanoh T, Akamatsu H, Kurisaka M. 1996. Role of cytokines, tyrosine kinase and protein kinase C on production of superoxide and induction of scavenging enzymes in human leukocytes. Clin. Immunol. Immunopathol. 79: 303–313. [ Links ]
Pallaoro A, Dulcic S, Matic–Skoko M, Kraljevic M, Jardas I. 2008. Biology of the salema Sarpa salpa (L. 1758) (Pisces, Sparidae) from the middle eastern Adriatic. J. Appl. Ichthyol. 24: 276–281. [ Links ]
Peirano A, Niccolai I, Mauro R, Bianchi CN. 2001. Seasonal grazing and food preference of herbivores in aPosidonia oceanica meadow. Sci. Mar. 367–374. [ Links ]
Quignard JP, Man Wai R, Vianet R. 1984. Les poissons de l'étang de Mauguio (Hérault, France): Inventaire, structure du peuplement, croissance et polymorphisme des tailles. Vie Milieu 34: 173–183. [ Links ]
Raikhlin–Eisenkraft B, Finkelstein Y, Spanier E. 1988. Ciguatera–like poisoning in the Mediterranean. Vet. Hum. Toxicol. 30: 352–353. [ Links ]
Rodríguez–Ariza A, Peinado JC, Pueyo C, López–Barea J. 1993. Biochemical indicators of oxidative stress in fish from polluted littoral areas. Can. J. Fish Aquat. Sci. 50: 2568–2573. [ Links ]
Romero J. 1989. Epífitos de las hojas de Posidonia oceanica: Variaciones estacionales y batimétricas de biomasa en la pradera de las islas Medes (Girona). Oecol. Aquat. 9: 19–25. [ Links ]
Sayeed I, Parvaez S, Pandey S, Bin–Hafeez B, Haque R, Raisuddin S. 2003. Oxidative stress biomarkers of exposure to deltamethrin in a freshwater fish, Channa punctatus Bloch. Ecotoxicol. Environ. Saf. 56: 295–301. [ Links ]
Shi HH, Sui YX, Wang XR, Luo Y, Ji LL. 2005. Hydroxyl radical production and oxidative damage induced by cadmium and naphthalene in liver of Carassius auratus. Comp. Biochem. Physiol. C 140: 115–121. [ Links ]
Smayda TJ. 1997. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanogr. 42: 1137–1153. [ Links ]
Steidinger KA, Baden DG. 1984. Toxic marine dinoflagellates. In: Spector DL (ed.), Dinoflagellates. Academic Press, Orlando, pp. 201–249. [ Links ]
Sureda A, Batle JM, Tauler P, Cases N, Aguilo A, Tur JA, Pons A. 2004. Neutrophil tolerance to oxidative stress induced by hypoxia/reoxygenation. Free Radic. Res. 38: 1003–1009. [ Links ]
Sureda A, Box A, Ensenat M, Alou E, Tauler P, Deudero S, Pons A. 2006. Enzymatic antioxidant response of a labrid fish (Coris julis) liver to environmental caulerpenyne. Comp. Biochem. Physiol. C 144: 191–196. [ Links ]
Tindall DR, Miller DM, Bomber JW. 1989. Culture and toxicity of dinoflagellates from ciguatera endemic regions of the word. Toxicon 27: 8–86. [ Links ]
Tomas CR, Throndsen J, Heimdal BR. 1993. Marine Phytoplankton: A Guide to Naked Flagellates and Coccolithophorids. Academic Press, 263 pp. [ Links ]
Tomas CR (ed.), Hasle GR, Syvertsen EE, Steidinger AK, Tangen K. 1996. Identifying Marine Diatoms and Dinoflagellates. Academic Press, 598 pp. [ Links ]
Tregouboff G, Rose M. 1957. Manuel de Planctonologie Méditerranéenne. Vol. II, CNRS, Paris, 587 pp. [ Links ]
Turki S, Harzallah A, Sammari C. 2006. Occurrence of harmful dinoflagellates in two different Tunisian ecosystems: The Lake of Bizerte and the Gulf of Gabes. Cah. Biol. Mar. 47:1–7. [ Links ]
Utermöhl H. 1958. Zur Vervollkommung der quantitativen Phytoplankton–Methodik. Mitteilungen der Internationalen Vereinigung für Theoretische und Angewandte Limnologie 9: 1–38. [ Links ]
Vaillant V, Caumes E, De Valk H, Mesnage V, Griffon AM. 2001. Intoxication alimentaire à la ciguatera: Savoir l'évoquer même en l'absence de voyage. Bull. Epidémiol. Hebd. 38. [ Links ]
Velimirov B. 1984. Grazing of Sarpa salpa L. on Posidonia oceanica and utilization of soluble compounds. In: Boudouresque CF, Jeudy de Grissac A, Olivier J (eds.), International Workshop on Posidonia oceanica Beds. GIS Posidonie Marseille, France, pp. 381–387. [ Links ]
Williams SL, Ruckelshaus MH. 1993. Effects of nitrogen availability and herbivory on eelgrass (Zostera marina)and epiphytes. Ecology 74: 904–918. [ Links ]
Wu JY, Zheng L, Wang JH. 2005. Contamination of shellfish from Shanghai seafood markets with paralytic shellfish poisoning and diarrhetic shellfish poisoning toxins determined by mouse bioassay and HPLC. Food Addit. Contam. 22: 647–651. [ Links ]
Zairi M, Rouis MJ. 1999. Impacts environnementaux du stockage du phosphogypse à Sfax (Tunisie). Bull. Lab. Ponts Chaussées 219: 29–40. [ Links ]
No hay comentarios:
Publicar un comentario